|
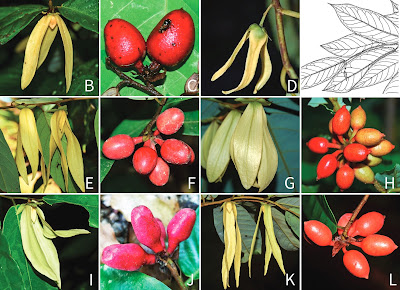
| A–C. ส่าเหล้า Friesodielsia brevistipitata Leerat. D. บุหงาเขาหลวง F. khaoluangensis Leerat. & Aongyong. E–F. บุหงาแต่งงาน F. longipetala Leerat. & Chalermglin. G–H. บุหงาสาวิตรี F. macrosepala Leerat. & Aongyong. I–J. บุหงาเซิงพะงัน F. phanganensis Leerat. K–L. บุหงาเซิงสงขลา F. songkhlaensis Leerat. in Leeratiwong, Chalermglin, ... et Johnson, 2021. |
Abstract
Six new species of Friesodielsia (Annonaceae) are described and illustrated, seven species are documented for Thailand for the first time, and Oxymitra fornicata var. glabra is raised to species rank as Friesodielsia andamanica. These additions bring the species diversity of Friesodielsia in Thailand to 16 species. A key to the Thai species is provided.
Keywords: Plant diversity, Southeast Asia, taxonomy
 |
| Photographs of new Friesodielsia species. A–C. Friesodielsia brevistipitata Leerat. A. leaf base showing marginal glands. B. flower. C. monocarps. D. flower of F. khaoluangensis Leerat. & Aongyong. E–F. F. longipetala Leerat. & Chalermglin. E. flowers, F. monocarps. G–H. F. macrosepala Leerat. & Aongyong. G. flowers. H. monocarps. I–J. F. phanganensis Leerat. I. flower. J. monocarps. K–L. F. songkhlaensis Leerat. K. flowers. L. monocarps. Photographs by C. Leeratiwong (A–C, E–F, I &K–L), K. Aongyong (G & H) and P. Khiriwong (J). |
Friesodielsia brevistipitata Leerat., sp. nov.
Friesodielsia brevistipitata is most similar to F. desmoides (Craib) Steenis and F. songkhlaensis Leerat. It differs from F. desmoides in having appressed rather than erect hairs on the abaxial leaf surface, sepals 5–10 mm versus 4–5 mm long, outer petals 35–60 mm versus (19–)27–31 mm long and obtuse versus acute at the apex, and monocarp stipes 3–5 mm versus 7–10.5 mm long. From F. songkhlaensis it may be distinguished by the shorter sepals that are rounded to an obtuse or slightly acute apex and again by the short monocarp stipes.
Etymology.— Named for the short stipes of the monocarps, among the shortest in the entire genus.
Vernacular.— Sa lao (ส่าเหล้า)(Krabi).
Friesodielsia khaoluangensis Leerat. & Aongyong, sp. nov.
Friesodielsia khaoluangensis resembles F. alpina (J.Sinclair) Steenis and F. borneensis (Miq.) Steenis in its narrow flowers and narrow leaves. From F. alpina it differs in the twigs and leaves appressed-pubescent rather than glabrous, larger leaves (13–22 × 3.8–7.5 cm versus 8.5–13 × 3–4 cm) but with only 9–12 secondary veins per side versus 12–14 per side, the pedicels ca 10 mm long versus 25–27 mm long, and the outer petals linear-lanceolate, 23–25 mm long, and appressed-pubescent versus lanceolate, ca 15 mm long, and glabrous. From F. borneensis it differs in the leaves not glaucous abaxially and with petioles 8–17 mm long instead of glaucous abaxially with petioles 4.5–8 mm long, the pedicellar bract lanceolate rather than linear, and petals 23–25 mm long rather than 52–59 mm long.
Etymology.— The species is named for the type locality.
Vernacular.— Bu nga khao luang (บุหงาเขาหลวง) (General),
sao yut (สาวหยุด) (Nakhon Si Thammarat).
Friesodielsia longipetala Leerat. & Chalermglin, sp. nov.
Friesodielsia longipetala is distinguished from F. phanganensis and F. songkhlaensis by its sub-coriaceous to coriaceous leaves, outer petals (50–)80–140 mm long, inner petals (11–)15–25 mm long, generally longer pedicels (up to 12 mm long), and monocarps only sparsely pubescent.
Etymology.— Named for the greater length of both the outer and inner petals compared with those of its congeners.
Vernacular.— Bu nga taeng ngan (บุหงาแต่งงาน)(General),
sao yut (สาวหยุด)(Satun).
Friesodielsia macrosepala Leerat. & Aongyong, sp. nov.
Friesodielsia macrosepala is similar to F. unonifolia but differs in having twigs covered by persistent mixed spreading and appressed hairs, sepals (7–)11–17 × (5–)9–11 mm, and outer petals 50–80 × 11–22 (–32) mm, with a length:width ratio ofca 4:1. Friesodielsia unonifolia, in contrast, has twigs initially covered with appressed hairs but soon glabrate, sepals 7–9 × 5.5–7 mm long, and outer petals 18–35 (–40) × 9.5–12 mm with a length:width ratio of ca 2.5:1.
Etymology.— The sepals are among the largest in the entire genus.
Vernacular.— Bu nga sawitri (บุหงาสาวิตรี)(Nakhon Si Thammarat),
sao yut (สาวหยุด)(Nakhon Si Thammarat)
Friesodielsia phanganensis Leerat., sp. nov.
Friesodielsia phanganensis is distinguished from F. longipetala by the chartaceous leaves, oblanceolate to elliptic-oblanceolate outer petals 50–70 mm long, inner petals 11–15 mm long, and monocarps densely pubescent with shorter stipes 3–5 mm long.
Etymology.— The species is named for the type locality.
Vernacular.— Bu nga soeang phangan (บุหงาเซิงพะงัน)(General).
Friesodielsia songkhlaensis Leerat., sp. nov.
Friesodielsia songkhlaensis is distinguished from F. longipetala by the chartaceous leaves, pedicels 3–5 mm long with the bract exceeding the pedicel inlength, sepals tapering to a blunt-acuminate apex, outerpetals 6–12 mm wide, inner petals 8–10 mm long, and more densely pubescent monocarps.
Etymology.— Named for the Thai province in which it occurs.
Vernacular.— Bu nga soeng songkhla (บุหงาเซิงสงขลา)(General).
 |
| A–B. Friesodielsia affinis (Hook.f. & Thomson) D.Das. A. flowers. B. monocarps. C. immature fruit of F. glabra (Ridl.) D.M.Johnson. D–E. F. calycina (King) Steenis. D. open flower and bud. E. monocarps. F. flower of F. discolor (Craib) D.Das. G–H. F. glauca (Hook.f. & Thomson) Steenis. G. flower. H. monocarps. I–J. F. kingii (J.Sinclair) Steenis. I. flower. J. monocarps. K. flower of F. latifolia (Hook.f. & Thomson) Steenis. L. flower of F. unonifolia (A.DC.) Steenis. Photographs by C. Leeratiwong (A–C, E, H & K), A. Baka (D, G, I & J), P. Chalermglin (F) and A. Rungrueng (L). |
Friesodielsia species newly reported for the Flora of Thailand.
รายงานพบใหม่ในประเทศไทย
1. Friesodielsia affinis (Hook.f. & Thomson) D.Das (เครือเหนียว)
2. Friesodielsia calycina (King) Steenis (บุหงาดาริกา)
3. Friesodielsia filipes (Hook.f. & Thomson) Steenis (บุหงาก้านดอกยาว)
4. Friesodielsia glauca (Hook.f. & Thomson) Steenis (บุหงานวลแป้ง)
5. Friesodielsia kingii (J.Sinclair) Steenis (บุหงากลีบใหญ่)
6. Friesodielsia latifolia (Hook.f. & Thomson) Steenis (บุหงาใบกว้าง)
7. Friesodielsia unonifolia (A.DC.) Steenis (บุหงากลีบหนา)
ปรับปรุงชื่อ ตามข้อมูลทางอนุกรมวิธาน
*Friesodielsia glabra (Ridl.) D.M.Johnson (บุหงาตะรุเตา)
Charan Leeratiwong, Piya Chalermglin, Jiratthi Satthaporn, Kithisak Aongyong and David M Johnson. 2021. New Species and New Records for the Climber Genus Friesodielsia (Annonaceae) in the Flora of Thailand. Thai Forest Bulletin (Botany). 49(2); 212-230. DOI: 10.20531/tfb.2021.49.2.09
ชนิดใหม่ในประเทศไทย
1. Friesodielsia brevistipitata Leerat., sp. nov. (ส่าเหล้า)
2. Friesodielsia khaoluangensis Leerat. & Aongyong, sp. nov. (บุหงาเขาหลวง)
3. Friesodielsia longipetala Leerat. & Chalermglin, sp. nov. (บุหงาแต่งงาน)
4. Friesodielsia macrosepala Leerat. & Aongyong, sp. nov. (บุหงาสาวิตรี)
5. Friesodielsia phanganensis Leerat., sp. nov. (บุหงาเซิงพะงัน)
6. Friesodielsia songkhlaensis Leerat., sp. nov. (บุหงาเซิงสงขลา)